
Source: http://tolweb.org/Terrestrial_Vertebrates/14952
Стегоцепхалианс: тетраподи и остали знаци кичмене вретенције
Мицхел Лаурин (Michel Laurin)

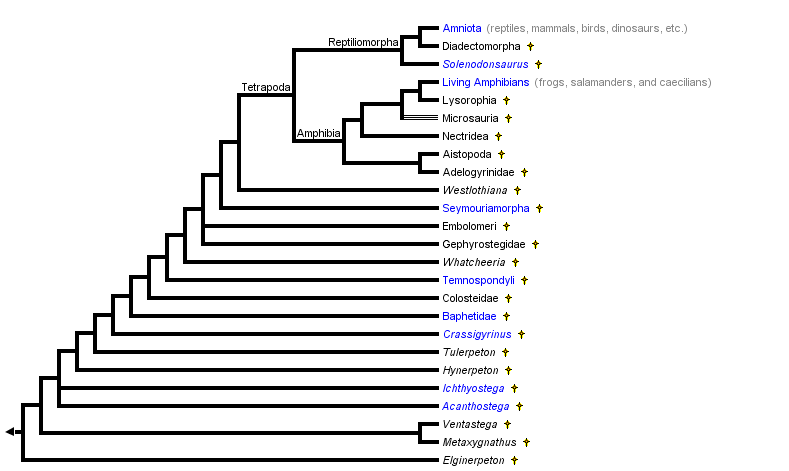

Измењено из Царролл (1995), Лаурин и Реисз (1997), и Лаурин (1998а-ц). Положај Вхатцхеериа прати Ломбард и Болт (1995). Положај слабо познатог девонског таксона (она у којима је неизвестан присуство цифара: Елгинерпетон, Метакигнатхус, Вентастега и Хинерпетон) прати Ахлберг (1998). Темноспондилс се често сматра се да ране водоземци. Међутим, приметити да у овом Пхилогени, нису уско повезани са лиссампхибианс.
На овој страници се говори о копненим клиторисима и другим кичмењачима који носе удке са цифрама, а не перфорама. Мишићни крак карактеристичан за ову класу углавном има добро дефинисане зглобове и цифре (прсте и прсте) и назива се хидридијум. Ова група обухвата око 21100 постојећих врста и вјероватно много већи број изумрлих врста. Тетраподи су само једна од неколико група кичмењачких лешева (погледајте одељак о класификацији испод).
Земаљски кљућари имају светску дистрибуцију. Најранији чланови ове групе били су умјерено велики (1-2,5 м дужине каросерије). Најстарији познати скелетни остаци копнених кичмењака пронађени су у Горњој Девонији Источне Гренландије (Цлацк, 1994). Присуство нижих до средњих девонијских путањи у Аустралији довело је до сугестија да је ова група можда настала у доњем девонском, прије најмање 400 милиона година (Варрен и сар., 1986), али цифре нису видљиве у овим утисцима, тако да ови путеви можда су оставили други саркоптери.
Највећа група земаљских кичмењака је Тетрапода (види одељак “Класификација копнених вретенчарија”, доле). Тетрапода значи “четири метра”, а група је била толико названа јер су његови чланови примитивно имали четири удове, за разлику од пераја. Овај таксон обухвата око 3000 постојећих врста водоземаца (жабе, саламандери и цаецилианс) и око 18100 постојећих врста амниота (сисара, гмизаваца и птица). Број изумрлих врста тетрапода је наравно непознат, али је око половине тренутно познатих врста тетрапода изумрло (Царролл, 1988).
Тетраподи настали су најкасније касније од Мисисипића (пре око 350 милиона година), од којих су познати најстарији познати рођаци живих водоземаца. У то вријеме морају бити присутни рођаци амниота, али до сада нису пронађени. Ранији извештај о фосилу (Смитхсон и сар., 1994; Царролл, 1995) као рани Миссиссиппиан амниоте или антхрацосаур (Вестлотхиана) вероватно је стем-тетрапод или рани амфибијанс (Лаурин, 1998а). Стегоцепхалианс (види одељак о класификацији испод за дефиницију ове групе) настао је најкасније од Горњег Девоније.
Тетраподи се крећу од 9,8 мм (у жабици Псиллопхрине дидацтила) до 30 м (у плавом киту) у укупној дужини. Они имају светску дистрибуцију и насељавају сва главна станишта. Већина су земаљски, али се неколико људи вратило у водено окружење у којем су живјели наши удаљени преци. Водени тетраподи укључују различите саламандере (сирениде, криптобранхиде, протеиде итд.), Жабе (пипиде), неке цаецилиане (тифлонектиде), кожне корњаче, морске змије, пиннипеде (печат и морусе) и китове. Неки тетраподи су способни да летају (птице и слепи мишеви), док остали клизају, као што су летеће веверице, дермоптерани (понекад се називају “летећи лемури”, иако нису примати) и летећи змај (Драцо воланс).
Страница Историја живот стегоцепхалианс садржи информације о овој комплексној теми.
Главни орган дисање већине стегоцепхалианс је плућа, али други респираторних органа постоје у многим групама. Више детаља је доступно на дах у стегоцепхалианс страници.
Многи стегоцепхалианс имају бубањ за особе високог фреквенцију, клима-борне звукове, а бочне линије орган наћи у многим водених водоземаца. За више информација, погледајте расправа у стегоцепхалианс страници.
Карактеристике
Стегоцефалци имају обиман фосилни запис (Царролл, 1988). Филогенетске студије су откриле неколико изведених карактеристика (синапоморфеја) стегоцефалијана:
- Губитак неколико кранијалних костију. У пандерицхтхиидс (групи сарцоптеригианс највише уско повезана са стегоцепхалианс), лобања је круто повезан са раменог појаса неколико костима који су нестали почетком у еволуцији земаљских кичмењака. Губитак ових костију и дозволио појаву мобилног врата која омогућава глава да се помера у односу на гепек. Ово раздвајање омогућава да глава остане релативно стабилна док хода.
- Губитак од оперцулар кости које покривају комору гилл у коштаних риба. Оперцулум више није потребан у раним цхоанатес јер су изгубили унутрашње шкрге својих раних предака. Међутим, оперцулум можда су нестали пре него што интерни шкрге (Цоатес и клак, 1991).
- Смањење у нотоцхорд и крута кичме. Кичменог центра за остеолепиформс су танки и окружује нотоцхорд (круту шипку присутна у свим пјевачице и, изнимно која траје у човеку као и интервертебрал дискова) без цонстрицтинг у великој мери. У стегоцепхалианс је центра су дебели и они ограничавају на нотоцхорд. Посебне артицулатори површине (у зигапопхисес) повезати неуро лукова једни другима. Краћи нотоцхорд да не продужи у браинцасе. Нотоцхорд од остеолепиформс продужен до близини хипофизе.
- Четири мишићни удови са дискретним цифре (прстима и ногама). Еарли финнед сарцоптеригианс попут Еустхеноптерон имао меснате ламела са елементима хомологна хумеруса, радијуса, улна, интермедиум, улнаре, фемур, тибиа, фибула и фибуларе, али хомологије више дисталних удова елемената је неизвесна и контроверзна. То је тренутно нејасно да ли сегментираног, разгранате ендоскелетал радиалс у дисталном делу пераје на сарцоптеригианс попут Еустхеноптерон или Тиктаалик су хомологне са цифрама. Оба аргумента за (Шубин и др., 2006) и против (Лаурин, 2006) Ова хипотеза је представљен.
- Сакралних ребро повезује аксијално костур (кичме) на карличног појаса (Тхе Хип). Ово омогућава тежина тела тетраподс који се преноси на задњу удова. Није било коштани веза између карличног појаса на остеолепиформс и њихове аксијално скелета. Губитак дерма фин зрака (модификованих скале који подржавају пераја). То једноставно представља уклањање објекта који више није потребна, а можда чак су штетни на копну.
Ови знакови се није појавио истовремено и изненада. Најстарији познати стегоцепхалианс, попут Ицхтхиостега Ацантхостега Ацантхостега Ацантхостегаи, поседују междинни услови за неке од ових ликова и недостају им друге. На пример, Ицхтхиостега задржао субоперцулар, кост која је део оперцулар комплекса који покривали гилл комору остеолепиформс. Ацантхостега задржао аноцлеитхрум, што је један од елемената који су водили раменом на лобањи у остеолепиформс (Цоатес и Цлацк, 1991). Нотоцхорд оф Ицхтхиостега и проширио дубоко у браинцасе, а већину свог каудалног пршљенова недостајало зигапопхисес (Јарвик, 1952). Веза између сакралног ребра и карличног појаса у је и даље слабо дефинисана. На крају, како Ицхтхиостега и Ацантхостега задржати лепидотрицхиа у репу, што указује да су таксона још увек је имао репне пераје.
На претходној листи су укључени само скелетни знаци, јер су све најраније групе стегацефалија изумрле, а мекани анатомски карактери се могу проучавати само у постојећим таксономима. Следећи знаци се налазе у тетраподима, али не и код других екстремних кичмењака:
- Слој мртвих, напаљених ћелија које смањује испаравајући губитак воде. Овај слој је присутан у амниотес иу већини лиссампхибианс. Кератина помаже одржавању слојева липида који смањују губитак воде (Кератин сама има слабе хидроизолације својства).
- Добро развијен мишићни језик са жлезда. Међутим, неки лиссампхибианс имају само основну језик, као риба. Примарни језик је једноставно меснате пута на поду уста која недостаје својствена мишиће и са ограниченом покретљивошћу.
- Параштитне жлезде укључени у контролу нивоа калцијума у крви.
- Хардериан жлезда налази спреда на оку. Ова жлезда лучи уљана течност која подмазује око.
- Вомеронасал (Јацобсон) орган. Овај мирисни органа налази се у непца и вероватно користи да смрди храну у уста.
- Губитак унутрашњих шкрге. Спољни шкрге присутни у многим водених и ларви лиссампхибианс су нове структуре и нису хомологне са унутрашњим шкргама риба.
Тешко је одредити тачно када се појавила ова ликови јер нису сачувани у фосилима, осим по основу индиректних трагове о унутрашњим шкрге, а најближи познати рођаци тетраподс су изумрли. Међутим, ови ликови се не налазе у лунгфисхес (најближи постојали рођаци тетраподс). Ацантхостега, А Девон стегоцепхалиан, још увек је имао унутрашње шкрге (Цоатес и клак, 1991), и Ицхтхиостега можда их задржао превише, али ни један други стегоцепхалиан се зна да их је имао. Због тога, унутрашње шкрге су вероватно изгубљени рано у еволуцији стегоцепхалианс, у девона или Миссиссиппиан (пре око 360 милиона година), а не тетрапод икада имао унутрашње шкрге.
Класификација земаљске кичмењака
У прошлости су већина земаљских хоганата укључена у Тетрапода (Гаффнеи, 1979). Недавно је Тетрапода формално дефинисан као крунска група (Гаутхиер и сар., 1989). Круна група је клад који укључује последњи заједнички предак две или више постојећих таксона и свих његових потомака. У овом случају, Тетрапода је дефинисана као клада која укључује последњи заједнички предак лисампибијаца и амниота, и све његове потомке.
Према Гаутхиер ет ал. (1989), Тетрапода је укључивао најпознатије фосилне земаљске кичмењаке, јер се сматра да су темноспондили стем-група амфибија, док се сматрало да су емболомери, гефиростегиди и сеимуриаморфи уско повезани са амниотима него са лизампибијцима. Због тога је из Тетраподе искључено само неколико врло раних копнених кичмењака, као што су Ихтиостега и Ацантхостега.
Овдје предложена филогенија говори да темноспондили, емболомери, гефиростегиди и сеимуриаморфи нису дио крунске групе. Ако је то тачно, ови таксони нису тетраподи, а порекло “тетраподног удова” предстоји порекло Тетрапода. У првим верзијама ове странице, сви саркоптерији који су носили цифре једноставно су назвали земаљски кичмењаки, јер није предложена формална филогенетска таксономија ове целине. Таква таксономија је недавно објављена (Лаурин, 1998а). Таксон Стегоцепхали (који је дуго укључивао све кичмене четворице, са изузетком лизампабијаца и амниота) дефинисани су као сви који су уско повезани са Темноспондили него са Пандерицхтхисом (најближи релатив тетраподова за које се зна да су задржали парене плавусе). Због тога, Стегоцепхали укључује све таксоне који садрже цифре, а неколико (Елгинерпетон, Метакигнатхус, Вентастега, и Хинерпетон) који могу задржати упарене пераје. Супротно старој употреби овог термина, Стегоцепхали се сада односи на клад. Израз “стегоцепхалиан” биће коришћен у наставку, јер не преноси хипотетичку и вероватно нешто погрешну интерпретацију да су сви бродски вретенчари земаљски. Као што је објашњено у даљем тексту, најранији чланови те класе вјероватно су примитивно водени.
Напомена о чвоу означеном Амфибијом: Други ограничавају име Амфибија на потомке најновијег заједничког претка постојећих водоземаца (терминалски таксон Живети амфибије на овом стаблу). Аутор преферира употребу наведену на горе наведеном стаблу, а дефиниција Амфибије као и сви тетраподи који су блиско повезани са постојећим водоземцима него амниотима има историјски приоритет (Лаурин, 1998а), али друга употреба је прилично распрострањена и не може се занемарити. На овој страници, термин Амфибија се увек односи на чвор означен на горе наведеном стаблу, а постојећи амфибије се називају лизампибијцима.
Напомена о чвору обележеном Тетраподом: Други проширују употребу овог имена како би укључили све кичмењаче који поседују удове цифрама (Лаурин и Андерсон, 2004). Међутим, на овој страници, Тетрапода се увек односи на чвор означен на горе наведеном стаблу.
Дискусија о филогенетских односа
Филогенија стегоцефалијана је контроверзна. Скоро сви систематисти се слажу да постојећи амфибије формирају монофилетску групу (Лиссампхибиа) која је блиско повезана са амниотима, али порекло лизампибијаца и амниота је контроверзна. Лаурин анд Реисз (1997, 1999), Лаурин (1998а, б), Валлин и Лаурин (2004) сугеришу да су лепоспондили матична група водоземаца и да су диадектоморфи блиско повезани са амниотима него са лизампибијцима. Сходно томе, многе групе палеозојских стегоцефалијана, као што су темноспондили и сејмуриаморфи, нису уско повезане са амниотима или лизампаминима:

Међутим, већина ране студије и неколико недавно они су предложили да се лиссампхибианс изведени из темноспондилс, и то диадецтоморпхс, сеимоуриаморпхс, а емболомерес се ближе односе на амниотес него лиссампхибианс (Болт, 1969, Гаффнеи 1979;. Готје и др, 1988, 1989, Милнер, 1988, 1993; Панцхен и Смитхсон, 1988; Труеб и Цлоутиер, 1991; Ломбард и Сумида, 1992;. Рута ет ал, 2003а, б; Рута и Цоатес, 2007). Лепоспондилс су често гледа као на парапхилетиц група која је укључивала ране родбину темноспондилс и лиссампхибианс. Стога, најпознатије стегоцепхалианс се гледало као повезано за било лиссампхибианс или амниотес. Само Ицхтхиостега и други Девон таксона (Ацантхостега, Тулерпетон) нису веровали да се на једном од ова два главна еволуционих родова:

Неколико палеонтолога и даље предлажу полифиличко порекло постојећих водоземаца (Андерсон, 2007, 2008; Царролл, 2007; Андерсон и сар., 2008), са гимнофионима угнеженим у “лепоспондилима” и ануранима и уроделима угнеженим у темноспондилима. Ово подразумева да су гимнафионици блиско повезани са амниотима него анурансима и уроделима. Ова филогенија је некомпатибилна са свим објављеним молекуларним и морфолошким филогенезама, што указује на то да постојећи амфибије формирају клад који искључује амниоте (Лаурин, 2002). Осим тога, поновна процена кодирања главне носиве матрице показује да она у ствари подржава монофилетно порекло лисампибијаца међу “лепоспондилима” (Марјановић и Лаурин, 2009). Развијени подаци су такође предложени да подрже постојеће полифилије амфибије (Сцхоцх и Царролл, 2003; Фробисцх и сар., 2007), али ови аргументи су одбијени (Сцхоцх, 2006, Гермаин и Лаурин, 2009, Марјановић и Лаурин, 2009). Развојни подаци не разликују различите хипотезе о пореклу лиссампхибиана.
Најновија пхилогени заговара лиссампхибиан полипхили (Андерсон ет ал, 2008) делом заснована на је недавно описани темноспондил Геробатрацхус хоттони, која је у почетку интерпретирана као сестра-групу Батрацхиа, која је дубоко уметнуте у темноспондилс; Гимнопхиона је смештен у “лепоспондилс”. Закључак је делом Тај заснована на тумачењу различитих морфолошких карактера Геробатрацхус, Као што педицели зуба (а лиссампхибиан синапоморпхи), базална комуна (фузионисан дистални тарсалс 1 и 2; а батрацхиан синапоморпхи) и узак, бочно режији палатина (а салиентиан синапоморпхи). Међутим, присуство синапоморпхиес неколико уметнутих кладе, и други, много примитивнији знакова, као што је задржавање постпариетал, табеларно и супратемпорал, указује велико хомопласи. Тумачења морфологији Друге Геробатрацхус хоттони су Могуће и инкорпорација том том облику у матрици првом података ко је је подржала лиссампхибиан полипхили (МцГован, 2002) и даље подржава монопхилетиц порекло Лиссампхибиа унутар лепоспондилс (Марјановић и Лаурин, 2008б).
Неколико молекуларних студија предложило је врло ране датуме за почетак диверсификације лиссампхибиан-а, у распону од око 337 Ма (Зханг ет ал., 2005) до 362 Ма (Сан Мауро ет ал., 2005; Роелантс ет ал., 2007). Зханг ет ал. (2005) су такође сугерисали да је ово утицало на то да се лизампибијанци добијају из темноспондила, али Лее и Андерсон (2006) истичу да су такви датуми ране диверзификације били више компатибилни са полифилетичком хипотезом, у којој екстантни водоземци стварају клад који искључује амниоте, али проистичу из оба “лепоспондила” и темноспондила. Међутим, ново молекуларно и палеонтолошко давање диверзификације лисампибијаца сугерише много новију диверзификацију ове групе у пермијану (Марјановић и Лаурин, 2007). Израчунавање интервала поверења на прави стратиграфски распон Лиссампхибиа, засновано на даљем развоју најсофистицираније методе коју је развио Марсхалл (1997), такође сугерише пермско, монофилијско порекло групе (Марјановић и Лаурин, 2008а).
Додатне дискусија и детаљнији пхилогениес могу се наћи на Пхилогени на стегоцепхалианс странице.
Порекло стегоцепхалианс
Неколико сценарија предложено је да објасни порекло цхиридиум и зашто кичмењака одважио на копну. Теорија наћи у већини популарних књига је да аридним клима која је некада сматрало да су превладали у девона су приморани наше сарцоптеригиан прецима да пузи од сезонски сушења језера да се постигне веће, дубље тела воде (Ромер, 1933). Међутим, проблем са овом сценарију је да Девон више не гледа као да је био сезонски сува, и класичне Лате Девон формације, као што је Есцуминац у Квебеку, Канада, сада мислили да су Естуарине (дакле, не сезонски суша могла евентуално јављају, као што је у великој мери зависи од доба ниво мора). Пераја ендоскелетон од ребрастих тетраподоморпхс, недавно је студирао у тристицхоптерид Еустхеноптерон, Очигледно веома сунђерасти и појављује се лоше одговара да се одупре значајан механички стрес на дужи временски период, како би дошло у дугим копнених излета (Лаурин и др., 2007). Осим тога, новије студије о функционалној морфологије указују на то да су удови су еволуирали да би наши рани преци да хода на дну плитких језера или мочварама (Едвардс, 1989). Откриће прилично комплетних примерака у девонског цхоанате Ацантхостега (Цоатес и Цлацк, 1990) потврђује да њени удови су били слабо одговара да хода по копну (Зиммер, 1995). Претпостављен Присуство унутрашњих шкрга ин Ацантхостега такође потврђује да ова животиња још увек углавном водене (Цоатес и Цлацк, 1991). Изгледа да су се дуже него што се раније мислило Прелаз из водену на земаљској животној средини.
Рано еволутион оф тхе лимбс
Елементи хомологне са стилопод (бутне кости и десног) и зеугопод (радијуса, улне, тибије и фибуле) су одавно познато да се у тетраподоморпхс (нпр Грегори, 1911), али до недавно, мало се знало о времену настанка разни ликови из тетрапод екстремитета. Много је постигнут напредак на овом пољу недавно. На пример, недавна хистологијска студија је показала да Еустхеноптерон фоорди упарени пераја поседовао истински дуге кости које расте у дужини и дебљини преко ендоцхондрал и периосталног осификације, респективно, као у тетраподс (Меуниер и Лаурин, у штампи). Боисверт ет ал. (2008) су показали да је дистални део грудног пераје на Пандерицхтхис, Један од највећих цровнвард финнед тетраподоморпхс, састоји се од неколико Радиалс, као што треба очекивати у облику уско везано за стегоцепхалианс. Изгледа да је погрешно преведено раније реконструкције који је представљао једну велику дистални тањир уместо неколико мањих Радиалс.
До недавно, најстарији унрефутабле докази о присуству цифара је представљао скелетних остатака из самог краја девона (Фаменниан). Четири Девон таксони условно укључена у дрвету изнетог су познати из фрагментарних остатака. Фрагментарних природа остатака ових родова спречава самоуверен процену њиховим афинитетима, иако Цоатес (1996) и Ахлберг (1998) су предложили да се ближе односе на тетраподс него пандерицхтхиидс (најближи познати рођаци тетраподс познато да имају задржава упареним пераја). Ови Енигматиц така спадају Елгинерпетон (Ахлберг, 1995), Вентастега (Ахлберг ет ал., 1994), Метакигнатхус (Цампбелл и Белл, 1977), анд Хинерпетон (Даесцхлер ет ал., 1994). Њихова апендикса скелет је слабо (или не) познате, и они су искључени из најмањег клада која укључује лимбед кичмењака. Зато, можда су задржане пераја (Лаурин ет ал., 2000). Међутим, недавно откриће Средњи Девон (Еифелиан) стаза (Ниедзвиедзки и др., 2010) у напуштеном каменолому Зацхеłмие (Пољска) чини вероватније да облици као што су Елгинерпетон поседовао цифара. Ниедзвиедзки ет ал. (2010) даље указују да је ова стаза подразумева да Девон стегоцепхалианс имају веома лошу фосилни запис, у мери у којој је то подразумева (по њиховом мишљењу) присуство шест дугих духова родова пружају од Миддле преко касног девона у стегоцепхалианс, поред додатним духова родах у блиско повезаних тетраподоморпхс, попут Пандерицхтхи, Тиктаалик, Елпистостеге и Ливониа (види слику, део А). Међутим, ова тврдња може бити преувеличана у тој мери да тачан систематски положај Зацхеłмие трацкмакер је непознат. Она замислити може представљати више базалну форму, искључен из најмањих клада која укључује Елгинерпетон, Вентастега и више цровнвард облика, а то би подразумевало јединствени стегоцепхалиан духа лозу пружа од Миддле преко касног девона, поред неколико духова линеагес ин финнед тетраподоморпхс (Лаурин, 2010; фигура, део В).
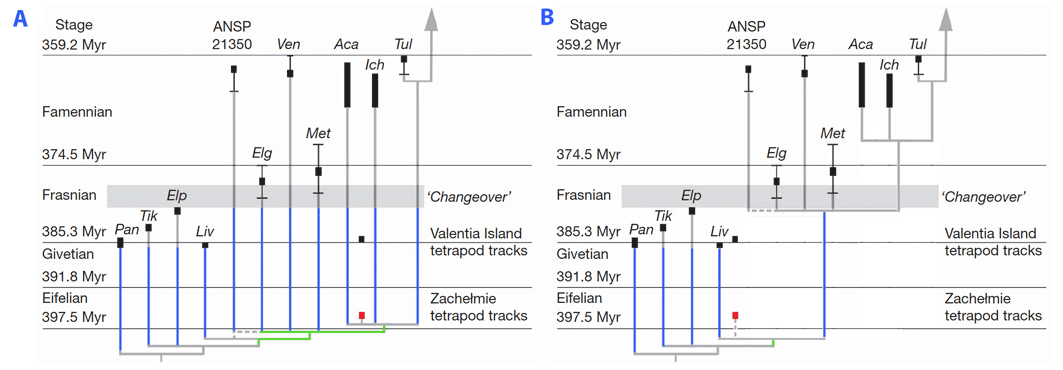
Могуће филогенетске позиције пољског средње девонијског стаза. А, позиција коју су предложили Ниедзвиедзки и сар. (2010). В, позиције су предложиле (у тексту) Лаурин (2010). С обзиром на расположиве доказе, ништа се не може одбити. Линије духова су плаве. Гране су одмах под могућом позицијом аутопута у зеленој боји. Део А је преусмерен са слике 5b Ниедзвиедзки и сар. (2010).
Присуство пет прстију је генерално верује да примитиван за тетраподс и да је био највећи број цифара које су наши рани преци имали. Пет цифре су вероватно били присутни у најранијим тетраподс, али не у најранијим стегоцепхалианс. Недавна открића су показала да су најранији стегоцепхалианс има више бројева; Ацантхостега имао осам прстију, Ицхтхиостега имао седам прстију, а Тулерпетон имао шест прстију (Лебедев, 1986; Гоулд, 1991). Ови “екстра” цифре су брзо изгубили, међутим, јер нико после Девон стегоцепхалиан је познато да је имао више од пет цифара.
Коутс (1991, 1996) је предложио да Пентадацтил услов појавила два пута (бар у цетвртом екстремитета), јер у његовим предложеним пхилогениес је Девон полидацтилоус стегоцепхалиан Тулерпетон се веровало да се ближе односе на амниотес него лиссампхибианс. Због тога, он је тврдио да је смањење од полидацтил до Пентадацтил стања дошло цонвергентли у антхрацосаурс (амниотес и њихових изумрлих рођака) и водоземаца (иако Пентадацтил стање у водоземаца је знао само из неколико узорака; већина лиссампхибианс имају само четири цифре у Манус). Међутим, реаналисис ових података, и уградња Тулерпетон у матрицу података која обухвата више така сугерише да Пентадацтил услов појавила само једном, и добро пред разилазе водоземаца и антхрацосаурс (Лаурин, 1998ц). Овај закључак произилази из постављања Тулерпетон у више базалној позицију у филогенија, ван клада који садржи све пост-ДеВониан (и све Пентадацтил) таксона.
Освајање земљишта од кичмењака
Први лимбед кичмењака, попут Ацантхостега и Ицхтхиостега, су очигледно водене таксона (Цлацк, 2002) јер изгледа да су задржали интерне шкрге, као што је показано жлебове на цератобранцхиалс које можда смештене аферентне шкржни артерије. Такође задржала постбранцхиал ламине о цлеитхрум, које можда омеђена су шкржни комору позади (Цоатес и Цлацк, 1991). Поред тога, они задржали лепидотрицхиа у каудалном пераје и добро развијеном бочне линије органа, као што је приказано каналима за свој кефалним порцији.
Унутрашње шкрге можда су рано расле у Царбонифероус и нема трагове лепидотрицхиа код стегоцепхалианс након Девоније, а то може указивати на нешто мање водени животни стил. Међутим, орган бочне линије је присутан код већине карбонских и многих пермијских стегоцефалијана, што указује на то да су многе врсте још увијек првенствено водене. Одсуство жлебова за орган бочне линије није поуздан критеријум за закључивање копненог начина живота јер орган може бити присутан без остављања трагова на скелету. Будући да је облик тела раних стегацефалија прилично конзервативан, водене врсте се не морају нужно драстично разликовати од терестричких у облику тијела (Лаурин, 2008). Тако су проучавани додатни критеријуми, као што је микроанатомија костију, да би се утврдило када су кичмењаче постале копнене (Лаурин и сар., 2004, Гермаин и Лаурин, 2005). Прелиминарни резултати указују на то да се појавио копенски начин живота код стегоцефалија у раном карбонизму (Крилофф и сар., 2008), али ће се морати проучити још много врста како би се дали поуздани резултати.
Концентрација атмосферског кисеоника је вероватно имала улогу у омогућавању кичмењака да се преселе на копно. Први, водени стегоцефалијани су се појавили док је атмосферски кисеоник био присутан у ниској концентрацији. Ромеров јаз, период у коме је познато врло мало стегацефалија, представља завршну фазу ове епизоде ниске концентрације кисеоника. Убрзо након што је концентрација атмосферског кисеоника постигла и чак превазишла садашњи ниво, стегоцефалије су спектакуларно еволуционо зрачење, у које су укључени и први копнени кичмени суплементи. То еволуционо зрачење је паралеловано код артропода. Тако, Вард ет ал. (2006) сугеришу да је концентрација кисеоника покренула или олакшала террестриализацију код артропода и кичмењака и да је концентрација кисеоника од око 20% у атмосфери доњи праг који је омогућио прелазак са жилног удисања у воду на ваздушно дисање код кичмењака и членоножаца.
Оригинал станиште стегоцепхалианс
Дуго се претпостављало да су први стегоцепхалианс биле слатке воде облици, као што су очуваних водоземаца и дипноанс (Ромер, 1933). Међутим, много истраживања од 1980. па надаље је показало да су ране сарцоптеригианс (укључујући дипноанс) су најчешће морски облици (Јанвиер, 1996). Слично, неколико класичних Пермо-карбон локалитети који су донели стегоцепхалианс, и који су дуго сматрана (барем кичмењака палеонтолога) представљају слатководних окружења недавно показано да покаже морског утицај (Лаурин и Солер-Гијон, 2006, 2010; Сцхултзе , 2009). Чини се да су многи од првих стегоцепхалианс су приморске облици, као што је приказано на Блиском девонским трацкваис, који су оставили у интертидал или лагоонал окружењу (Ниедзвиедзки и др., 2010), и јасно морски станиште Тулерпертон (Лебедев 1986). Међутим, неке од својих блиских рођака, као што је Тиктаалик (Даесцхлер ет ал., 2006), као и неких ранијих стегоцепхалианс, као што су Ицхтхиостега и Ацантхостега (Цлацк, 2002), да се интерпретира као слатководних облика. Врло је вероватно да стегоцепхалианс насељен слане, слатководних и копнених окружења крајем палеозоика. Изгледа да буде аутапоморпи од Лиссампхибиа (Лаурин и Солер-Гијон, 2010) је нетрпељивост у сланој води која карактерише већину лиссампхибианс.
Други називи за земаљско кичмењака
Стегоцепхалианс
Референце
Ahlberg P. E. 1995. Elginerpeton pancheni and the earliest tetrapod clade. Nature 373: 420-425.
Ahlberg P. E. 1998. Postcranial stem tetrapod remains from the Devonian of Scat Craig, Morayshire, Scotland. Zoological Journal of the Linnean Society 122: 99-141.
Ahlberg P. E., E. Luksevics, & O. Lebedev. 1994. The first tetrapod finds from the Devonian (Upper Famennian) of Latvia. Philosophical Transactions of the Royal Society B 343: 303-328.
Anderson, J. S. 2007 Incorporating ontogeny into the matrix: a phylogenetic evaluation of developmental evidence for the origin of modern amphibians. In: Major transition in vertebrate evolution, (Anderson, J. S. & Sues, H.-D., eds.). pp. 182-227. Indiana University Press, Bloomington.
Anderson J. S. 2008. The origin(s) of modern amphibians. Evolutionary Biology 35: 231–247.
Anderson J. S., R. R. Reisz, D. Scott, N. B. Fröbisch, & S. S. Sumida. 2008. A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders. Nature 453: 515–518.
Boisvert C. A., E. Mark-Kurik, & P. E. Ahlberg. 2008. The pectoral fin of Panderichthys and the origin of digits. Nature 456: 636–638.
Bolt, J. R. 1969. Lissamphibian Origins: Possible Protolissamphibian from the Lower Permian of Oklahoma. Science 166: 888-891.
Campbell K. S. W. & M. W. Bell. 1977. A primitive amphibian from the Late Devonian of New South Wales. Alcheringa 1: 369381.
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution. New York: W. H. Freeman and Company.
Carroll, R. L. 1995. Problems of the phylogenetic analysis of Paleozoic choanates. Bulletin du Muséum national d’Histoire naturelle de Paris 4ème série 17: 389-445.
Carroll, R. L. 2007. The Palaeozoic ancestry of salamanders, frogs and caecilians. Zool. J. Linn. Soc. 150: 1–140.
Clack, J. A. 1994. Earliest known tetrapod braincase and the evolution of the stapes and fenestra ovalis. Nature 369: 392-394.
Clack J. A. 2002. Gaining ground: the origin and evolution of tetrapods. Bloomington: Indiana University Press, 369 pp.
Coates M. 1991. New palaeontological contributions to limb ontogeny and phylogeny. In: J. R. Hinchcliffe (ed.) Developmental Patterning of the Vertebrate Limb 325-337. New York: Plenum Press.
Coates M. I. 1996. The Devonian tetrapod Acanthostega gunnari Jarvik: postcranial anatomy, basal tetrapod interrelationships and patterns of skeletal evolution. Transactions of the Royal Society of Edinburgh 87: 363-421.
Coates, M. I. & J. A. Clack. 1990. Polydactyly in the earliest known tetrapod limbs. Nature 347: 66-69.
Coates, M. I. & J. A. Clack. 1991. Fish-like gills and breathing in the earliest known tetrapod. Nature 352: 234-236.
Daeschler E. B., N. H. Shubin, K. S. Thomson, & W. W. Amaral. 1994. A Devonian tetrapod from North America. Science 265: 639-642.
Daeschler E. B., N. H. Shubin, & F. A. Jenkins, Jr. 2006. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440: 757-763.
Edwards, J. 1989. Two perspectives on the evolution of the tetrapod limb. The American Zoologist 29: 235-254.
Fröbisch N. B., R. L. Carroll, & R. M. Schoch. 2007. Limb ossification in the Paleozoic branchiosaurid Apateon (Temnospondyli) and the early evolution of preaxial dominance in tetrapod limb development. Evolution & Development 9: 69-75.
Gaffney, E. S. 1979. Tetrapod monophyly: a phylogenetic analysis. Bulletin of the Carnegie Museum of Natural History 13: 92-105.
Gauthier, J., A. G. Kluge, & T. Rowe. 1988. The early evolution of the Amniota. In M. J. Benton (ed.) The phylogeny and classification of the tetrapods, Volume 1: amphibians, reptiles, birds: 103-155. Oxford: Clarendon Press.
Gauthier, J., D. C. Cannatella, K. De Queiroz, A. G. Kluge, & T. Rowe. 1989. Tetrapod phylogeny. In B. Fernholm, K. Bremer, and H. Jornvall (eds.) The Hierarchy of Life: 337-353. New York: Elsevier Science Publishers B. V. (Biomedical Division).
Germain D. & M. Laurin. 2005. Microanatomy of the radius and lifestyle in amniotes (Vertebrata, Tetrapoda). Zoologica Scripta 34: 335-350.
Germain D. & M. Laurin. 2009. Evolution of ossification sequences in salamanders and urodele origins assessed through event-pairing and new methods. Evolution & Development 11: 170–190.
Gould, S. J. Eight (or fewer) little piggies. Natural History, January 1991, 22-29.
Gregory W. K. 1911. The limbs of Eryops and the origin of paired limbs from fins. Science 33: 848-849.
Janvier P. 1996. Early vertebrates. Oxford monographs on geology and geophysics, Oxford: Oxford University Press, 393 pp.
Jarvik, E. 1952. On the fish-like tail in the ichthyostegid stegocephalians with descriptions of a new stegocephalian and a new crossopterygian from the Upper Devonian of East Greenland. Meddelelser om Grønland 114: 1-90.
Kriloff A., D. Germain, A. Canoville, P. Vincent, M. Sache, & M. Laurin. 2008. Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference. Journal of Evolutionary Biology 21: 807–826.
Laurin M. 1998a. The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I-systematics, middle ear evolution, and jaw suspension. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: 1-42.
Laurin M. 1998b. The importance of global parsimony and historical bias in understanding tetrapod evolution. Part II-vertebral centrum, costal ventilation, and paedomorphosis. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: 99-114.
Laurin M. 1998c. A reevaluation of the origin of pentadactyly. Evolution 52: 1476-1482.
Laurin, M. 2002. Tetrapod phylogeny, amphibian origins, and the definition of the name Tetrapoda. Syst. Biol. 51: 364-369.
Laurin, M. 2006. Scanty evidence and changing opinions about evolving appendages. Zoologica Scr. 35: 667-668.
Laurin M. 2008. Systématique, paléontologie et biologie évolutive moderne : l’exemple de la sortie des eaux des vertébrés. Collection Parcours LMD – Sciences de la vie et de la Terre, ed. J. Segarra. Paris: Ellipses, 176 pp.
Laurin M. 2010. How Vertebrates Left the Water. Translated by M. Laurin. Berkeley: University of California Press, xv + 199 pp.
Laurin M. & J. S. Anderson. 2004. Meaning of the name Tetrapoda in the scientific literature: an exchange. Systematic Biology 53: 68-80.
Laurin, M. & R. R. Reisz. 1997. A new perspective on tetrapod phylogeny. In S. Sumida and K. Martin (eds.) Amniotes Origins: Completing the Transition to Land: 9-59. London: Academic Press.
Laurin M. & R. R. Reisz. 1999. A new study of Solenodonsaurus janenschi, and a reconsideration of amniote origins and stegocephalian evolution. Canadian Journal of Earth Sciences 36: 1239-1255.
Laurin, M., Girondot, M. & de Ricqlès, A. 2000. Early tetrapod evolution. Trends Ecol. Evol. 15: 118-123.
Laurin M., M. Girondot, & M.-M. Loth. 2004. The evolution of long bone microanatomy and lifestyle in lissamphibians. Paleobiology 30: 589-613.
Laurin M. & R. Soler-Gijón. 2006. The oldest known stegocephalian (Sarcopterygii: Temnospondyli) from Spain. Journal of Vertebrate Paleontology 26: 284-299.
Laurin M. & R. Soler-Gijón. 2010. Osmotic tolerance and habitat of early stegocephalians: indirect evidence from parsimony, taphonomy, paleobiogeography, physiology and morphology. In: M. Vecoli and G. Clément (ed.) The Terrestrialization Process: Modelling Complex Interactions at the Biosphere-Geosphere Interface (339): 151–179. London: The Geological Society of London.
Lebedev, O. A. 1986. The first record of a Devonian tetrapod in the USSR. Doklady-Earth science sections 278: 220-222.
Lee, M. S. Y. & Anderson, J. S. 2006. Molecular clocks and the origin(s) of modern amphibians. Mol. Phyl. Evol. 40: 635-639.
Lombard, R. E. & J. R. Bolt. 1995. A new primitive tetrapod, Whatcheeria deltae, from the Lower Carboniferous of Iowa. Palaeontology 38: 471-494.
Lombard, R. E. & S. S. Sumida. 1992. Recent progress in understanding early tetrapods. The American Zoologist 32: 609-622.
Marjanović, D. & Laurin, M. 2007. Fossils, molecules, divergence times, and the origin of lissamphibians. Syst. Biol. 56: 369-388.
Marjanović D. & M. Laurin. 2008a. Assessing confidence intervals for stratigraphic ranges of higher taxa: the case of Lissamphibia. Acta Palaeontologica Polonica 53: 413–432.
Marjanović D. & M. Laurin. 2008b. A reevaluation of the evidence supporting an unorthodox hypothesis on the origin of extant amphibians. Contributions to Zoology 77: 149–199.
Marjanović D. & M. Laurin. 2009. The origin(s) of modern amphibians: a commentary. Evolutionary Biology 36: 336–338.
Marshall C. R. 1997. Confidence intervals on stratigraphic ranges with nonrandom distributions of fossil horizons. Paleobiology 23: 165-173.
McGowan, G. J. 2002. Albanerpetontid amphibians from the Lower Cretaceous of Spain and Italy: a description and reconsideration of their systematics. Zool. J. Linn. Soc. 135: 1-32.
Meunier F. J. & M. Laurin. in press. A microanatomical and histological study of the fin long bones of the Devonian sarcopterygian Eusthenopteron foordi. Acta Zoologica 19 pp., 3 figs, 1 table (Early View version available on the journal’s web site).
Milner, A. R. 1988. The relationships and origin of living amphibians. In M. J. Benton (ed.) The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds: 59-102. Oxford: Clarendon Press.
Milner, A. R. 1993. The Paleozoic relatives of lissamphibians. Herpetological Monographs 7: 8-27.
Niedzwiedzki G., P. Szrek, K. Narkiewicz, M. Narkiewicz, & P. E. Ahlberg. 2010. Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463: 43–48.
Nussbaum, R. A. 1977. Rhinatrematidae: A new family of caecilians (Amphibia: Gymnophiona). Occasional papers of the Museum of Zoology, University of Michigan 682: 1-30.
Panchen, A. L. & T. R. Smithson. 1988. The relationships of the earliest tetrapods. In M. J. Benton (ed.) The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds: 1-32. Oxford: Clarendon Press.
Roelants, K., Gower, D. J., Wilkinson, M., Loader, S. P., Biju, S. D., Guillaume, K., Moriau, L. & Bossuyt, F. 2007. Global patterns of diversification in the history of modern amphibians. Proc. Natl. Acad. Sci. U.S.A. 104: 887-892.
Romer, A. S. 1933. Vertebrate Paleontology. 1st ed., Chicago: University of Chicago Press.
Ruta, M. & Coates, M. I. 2007. Dates, nodes and character conflict: addressing the lissamphibian origin problem. J. Syst. Paleontol. 5: 69-122.
Ruta M., M. I. Coates, & D. D. L. Quicke. 2003. Early tetrapod relationships revisited. Biological Reviews of the Cambridge Philosophical Society 78: 251-345.
Ruta M., J. E. Jeffery, & M. I. Coates. 2003. A supertree of early tetrapods. Proceedings of the Royal Society of London, Series B 270: 2507-2516.
San Mauro, D., Vences, M., Alcobendas, M., Zardoya, R. & Meyer, A. 2005. Initial diversification of living amphibians predated the breakup of Pangaea. Am. Nat. 165: 590-599.
Schoch R. R. 2006. Skull ontogeny: developmental patterns of fishes conserved across major tetrapod clades. Evolution & Development 8: 524-536.
Schoch R. R. & R. L. Carroll. 2003. Ontogenetic evidence for the Paleozoic ancestry of salamanders. Evolution & Development 5: 314-324.
Schultze H.-P. 2009. Interpretation of marine and freshwater paleoenvironments in Permo–Carboniferous deposits. Palaeogeography, Palaeoclimatology, Palaeoecology 281: 126–136.
Shubin, N. H., Daeschler, E. B. & Jenkins, F. A., Jr. 2006. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 440: 764-771.
Smithson, T. R., R. L. Carroll, A. L. Panchen, & S. M. Andrews. 1994. Westlothiana lizziae from the Viséan of East Kirkton, West Lothian,Scotland, and the amniote stem. Transactions of the Royal Society of Edinburgh 84: 383-412.
Trueb, L. & R. Cloutier. 1991. A phylogenetic investigation of the inter- and intrarelationships of the Lissamphibia (Amphibia: Temnospondyli). In H.-P. Schultze and L. Trueb (eds.) Origins of the higher groups of tetrapods-Controversy and Consensus: 223-313. Ithaca: Comstock Publishing Associates.
Vallin G. & M. Laurin. 2004. Cranial morphology and affinities of Microbrachis, and a reappraisal of the phylogeny and lifestyle of the first amphibians. Journal of Vertebrate Paleontology 24: 56-72.
Ward P., C. Labandeira, M. Laurin, & R. Berner. 2006. Confirmation of Romer’s Gap as a low oxygen interval constraining the timing of initial arthropod and vertebrate terrestrialization. Proceedings of the National Academy of Sciences of the United States of America 103: 16818-16822.
Warren, A., R. Jupp, & B. Bolton 1986. Earliest tetrapod trackway. Alcheringa 10: 183-186.
Zhang, P., Zhou, H., Chen, Y.-Q., Liu, Y.-F. & Qu, L.-H. 2005. Mitogenomic perspectives on the origin and phylogeny of living amphibians. Syst. Biol. 54: 391-400.
Zimmer, C., Coming onto the land. Discover, June 1995, 118-127.
Информације на Интернету
За општи увод у тетраподс и њиховим блиским сродницима, пробајте Увод у Тетрапода (УЦМП, Беркли).
Наслов илустрациjе

| Научно име | Ацантхостега Гуннари Ицхтхиостега |
|---|---|
| Коментари | Тхе Девон земаљске цхоанатес Ацантхостега (дно) и Ицхтхиостега (на обали). |
| Упућивање | први пут објављена Дисцовер Магазине |
| Творац | Илустрација Алфред Камајиан |
| Спецификација стања | Фоссил – период: Девон |
| Ауторско право | © 1995 Alfred Kamajian (Алфреда Камајиан) |
О овоj страници
Желим да се захвалим г. Јохн Хутцхинсон, гђици Патрициа Лаи, и г. Маттхев Марлове који је уредио ову страницу. Ја сам задужен за др. Давид Маддисон који је пружио непроцењиву помоћ у форматирању ове странице, повезујући га са другим страницама на Дрвету живота, а чији су бројни предлози побољшали презентацију ове странице. Такође захваљујем Катји Сцхулз за провјеру мојих периодичних исправки пре него што их објавим.

Мусеум Натионал д’Хистоире Натурелле, Париз, Француска
Page copyright © 2011 Michel Laurin

Page: Tree of Life Terrestrial Vertebrates. Stegocephalians: Tetrapods and other digit-bearing vertebrates. Authored by Michel Laurin. The TEXT of this page is licensed under the Creative Commons Attribution License – Version 3.0. Note that images and other media featured on this page are each governed by their own license, and they may or may not be available for reuse. Click on an image or a media link to access the media data window, which provides the relevant licensing information. For the general terms and conditions of ToL material reuse and redistribution, please see the Tree of Life Copyright Policies.
Прво онлине 04 јануар 1996
Садржај је промењен 21. априла 2011
Laurin, Michel. 2011. Terrestrial Vertebrates. Stegocephalians: Tetrapods and other digit-bearing vertebrates. Version 21 April 2011.http://tolweb.org/Terrestrial_Vertebrates/14952/2011.04.21 in The Tree of Life Web Project, http://tolweb.org/